

Cáncer: más que un problema genético
Las mutaciones no son suficientes para explicar la proliferación descontrolada de células.

Probablemente, la mayoría de las personas tiene o tuvo un familiar o conocido con cáncer. Esta devastadora enfermedad se caracteriza por la proliferación descontrolada de células en un órgano o tejido específico, con capacidad de diseminarse a otras partes del cuerpo1, alterando su funcionamiento y, en muchos casos, provocando la muerte. Pese a los grandes esfuerzos y recursos invertidos en su erradicación, pocos tratamientos resultan realmente efectivos.
Quizás el fracaso radique en un punto fundamental: todavía no entendemos bien el cáncer.
La teoría de la mutación somática
Durante décadas, la ciencia entendió el cáncer como un problema genético. Alteraciones en genes que regulan el crecimiento, la proliferación o la muerte celular (oncogenes), o en genes supresores de tumores, provocan que las células se multipliquen sin control, formando tumores. Esta es la base de la teoría de la mutación somática.
Su origen se remonta a 1914, cuando el embriólogo Theodor Boveri propuso que el cáncer se genera por alteraciones en la cromatina de una célula. Fue la primera teoría coherente que vinculaba directamente las anomalías genéticas —específicamente cromosómicas— con el desarrollo del cáncer.
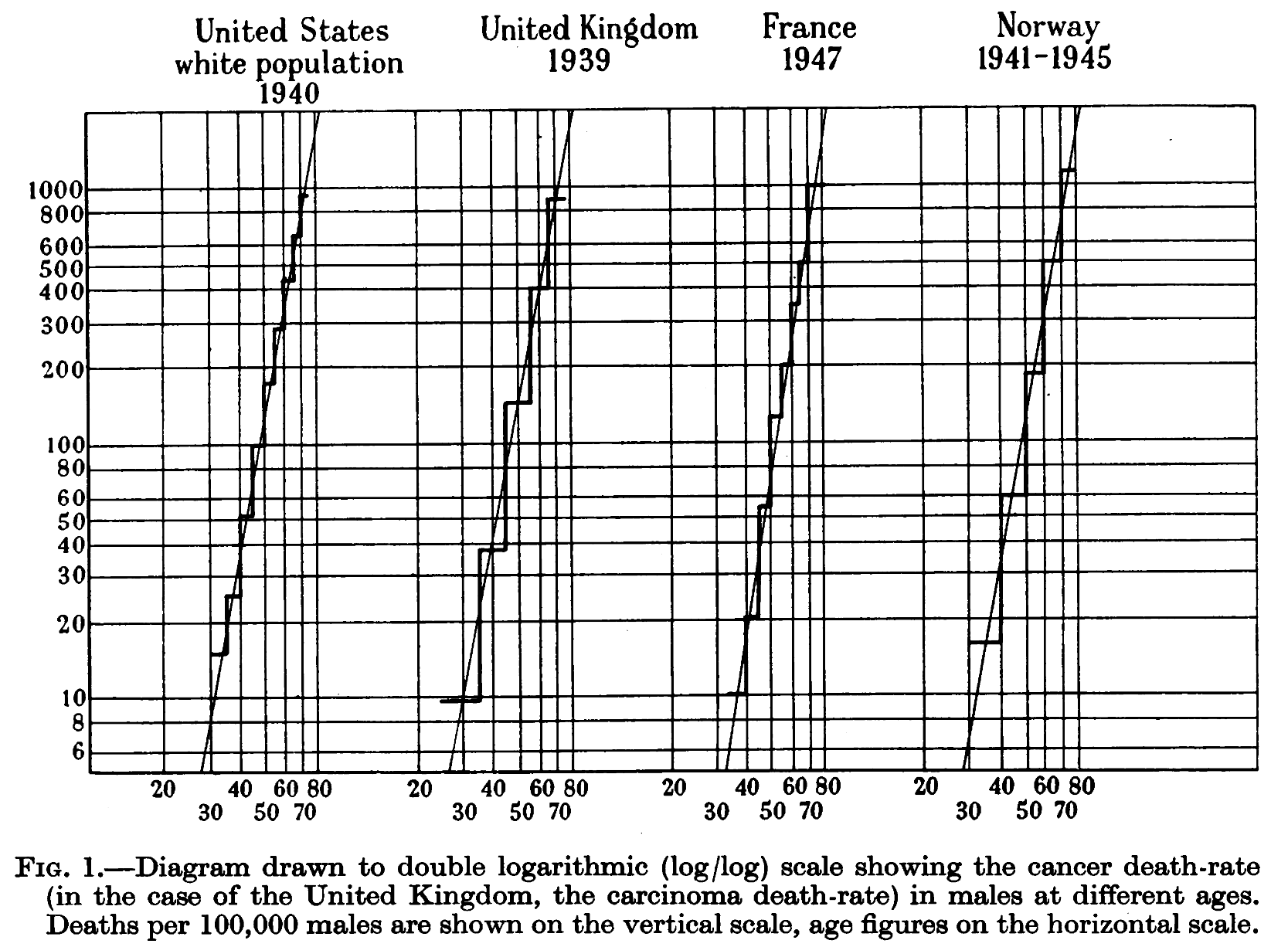
En la década de 1950, Karl Heinrich Bauer y Carl O. Nordling refinaron la teoría en términos de acumulación progresiva de mutaciones genéticas. Bauer se apoyó en los descubrimientos sobre agentes carcinógenos, como el alquitrán de hulla y los rayos X; mientras que Nordling observó una fuerte correlación entre la edad y la incidencia del cáncer.
Esta teoría aplica principios evolutivos al nivel celular. El cáncer sería un proceso evolutivo dinámico dentro del cuerpo: mutaciones aleatorias y selección natural actúan sobre las células somáticas2 en el microambiente tumoral, impulsando la progresión, heterogeneidad y resistencia terapéutica de los tumores.
La secuenciación genética desafía el paradigma
Tras el descubrimiento de la estructura del ADN por Watson y Crick en 1953 (basado en el trabajo de Rosalind Franklin y Maurice Wilkins), varios científicos desarrollaron técnicas para leer las secuencias genéticas. El método de terminación de cadena o “didesoxi”3, desarrollado en 1977 por el bioquímico británico Frederik Sanger, fue el más utilizado por su relativa eficiencia y robustez.
Gracias a estas técnicas, se identificaron mutaciones asociadas al cáncer, como las de los genes BRCA1 y BRCA24. Se diseñaron ratones modificados genéticamente para simular estos defectos, y la terapia dirigida prometía corregir las mutaciones específicas, lo que atrajo cuantiosos fondos.
Sin embargo, la misma tecnología de secuenciación que sustentó la teoría de la mutación somática del cáncer, también reveló sus limitaciones.
Con el tiempo, se detectaron numerosas mutaciones tanto en tejidos cancerosos como en tejidos normales, sin que se estableciera necesariamente una relación causal. Dos personas con la misma mutación podían tener resultados distintos: una desarrollaba cáncer, la otra no. Algunos tumores no presentaban mutaciones conductoras consistentes, mientras que tejidos normales albergaban mutaciones oncológicas sin convertirse en tumores. La heterogeneidad genética entre tumores del mismo tipo resultaba desconcertante.
Los genes no son los únicos culpables
La idea de que el cáncer no es una enfermedad exclusivamente genética, sino también una enfermedad del contexto celular —es decir, de los tejidos— no es nueva.
En los años 1970, la embrióloga Beatrice Mintz realizó experimentos innovadores que desafiaron la visión dominante sobre el cáncer y la biología del desarrollo. Su trabajo, centrado en la inoculación de células cancerosas en blastocistos de ratón, reveló mecanismos sorprendentes sobre cómo el entorno celular puede reprogramar células malignas.
Al inyectar células de teratocarcinoma (un tipo de tumor embrionario) en blastocistos de ratones sanos, los ratones quiméricos resultantes no desarrollaron tumores. Las células cancerosas, al interactuar con el entorno embrionario, se integraron en tejidos normales (hígado, piel, sistema nervioso) y funcionaron como células sanas. Esto demostró que las células cancerosas no son intrínsecamente "irreversibles"; su comportamiento depende del contexto.

A la inversa, células embrionarias normales introducidas en ambientes inadecuados —como el testículo o la cápsula renal— desarrollaban tumores, al perder las señales reguladoras del entorno uterino. Se formaban teratomas: tumores con tejidos desorganizados como pelo, dientes o músculo.
Estos hallazgos sentaron las bases de la teoría de organización de tejidos. En esta visión, las células cancerosas no actúan de forma autónoma. El microambiente —la matriz extracelular, las células vecinas y las señales químicas o mecánicas— puede activar o desactivar genes sin necesidad de mutaciones en el ADN. El cáncer se parecería más a un “ecosistema enfermo”, donde el suelo (entorno) es tan importante como la semilla (mutación).
El cáncer como un estado atávico
Las esponjas, uno de los animales más primitivos en el sentido evolutivo, conservan versiones ancestrales de muchos genes que, en humanos, están implicados en el cáncer. Esto sugiere que los oncogenes son antiguos y que originalmente cumplían funciones vitales en los primeros organismos multicelulares.
Sobre la base de estos hallazgos, el físico Paul Davies y el astrobiólogo Charles Lineweaver propusieron que el cáncer representa una reactivación de un estado biológico ancestral: un fenómeno atávico5. Los genes comúnmente alterados en el cáncer (oncogenes y genes supresores de tumores) son los más antiguos desde el punto de vista evolutivo. Estos controlaban funciones esenciales como proliferación, respuesta al estrés o metabolismo básico en los metazoos primitivos (Metazoa 1.0).
Con el tiempo, surgieron nuevas capas genéticas que suprimieron estas funciones primarias para dar lugar a organismos más organizados y cooperativos (Metazoa 2.0, 3.0...). Estas nuevas capas son responsables de la organización compleja de los tejidos, el desarrollo coordinado, la diferenciación celular especializada y la homeostasis (mantenimiento del equilibrio interno) en organismos multicelulares avanzados.
La proliferación autónoma y, hasta cierto punto, descontrolada de células parece ser un estado latente "por defecto" presente desde el último ancestro común universal de todos los organismos multicelulares. Por ello, cuando las capas reguladoras modernas fallan, las células pueden reactivar los antiguos programas genéticos: vuelven a ejecutar el “software” de Metazoa 1.0.
Atractores del cáncer
Llegado a este punto, vemos que el cáncer es una posibilidad inmanente a los metazoos y, por lo tanto, es menos adecuado para ser visto como "causado por algo", sino que es principalmente el desencadenamiento de algo latente. La carcinogénesis sería la entrada accidental en programas celulares antiguos.
Es así como surge el concepto de "atractores de cáncer", propuesto por Huang, Ernberg y Kauffman en 2009, el cual ofrece un nuevo marco para entender el cáncer no como un error genético, sino como una disfunción en la dinámica de redes reguladoras génicas (GRN)6.
En sistemas dinámicos, un atractor es un estado estable al que tiende un sistema. Cada tipo celular corresponde a un atractor estable en las GRN, definido por patrones específicos de expresión génica.
Las mutaciones pueden alterar el cableado de las GRN. En la mayoría de los casos, solo distorsionará suavemente la topografía del paisaje7. Hay un número limitado de formas en que un paisaje puede cambiar suavemente sin dejar de garantizar la viabilidad de la célula. Sin embargo, una mutación también podría facilitar la entrada accidental en un atractor de cáncer, impulsada por el ruido molecular o el estrés ambiental no mutagénico. En este caso, se puede admitir que las mutaciones causan cáncer.

Desde esta perspectiva, las células cancerosas no son solo células mutadas, sino células atrapadas en un estado disfuncional y estable. Este enfoque explica por qué no todas las mutaciones causan cáncer y por qué el microambiente y las señales epigenéticas juegan un rol crucial.
Los "atractores de cáncer" ofrecen un marco unificador para entender el cáncer como un error en la autoorganización celular, donde genética, epigenética y ambiente interactúan. Este enfoque podría transformar la oncología, priorizando terapias que restauren la regulación natural de las GRN en lugar de solo eliminar células mutadas.
Conclusión
En lugar de centrarse únicamente en las mutaciones genéticas, los científicos exploran cómo la regulación y cooperación entre los genes afecta los estados celulares. Los genes codifican directamente las proteínas, no los fenotipos, y no actúan solos. Los genes interactúan entre sí a través de las proteínas reguladoras (factores de transcripción), pequeñas moléculas de ARN y configuraciones cromosómicas para producir colectivamente un tipo celular.
Replantear el cáncer como un fenómeno de redes y contextos abre nuevas posibilidades terapéuticas: no solo corregir mutaciones, sino también restaurar el entorno que regula su expresión.
Referencia: Huang, S., Soto, A. M., & Sonnenschein, C. (2025). The end of the genetic paradigm of cancer. PLoS Biology, 23(3), e3003052.
Metástasis.
Son todas las células que forman el cuerpo de un organismo multicelular (como un humano), excepto las células germinales (los espermatozoides y los óvulos, que son las células reproductivas).
Se basa en la síntesis controlada de ADN in vitro utilizando una enzima (ADN polimerasa), un molde de ADN que se quiere secuenciar, y una mezcla de nucleótidos normales (dNTPs) y una pequeña cantidad de nucleótidos modificados llamados didesoxinucleótidos (ddNTPs). Esto genera una colección de fragmentos de ADN de diferentes longitudes, cada uno terminando en el ddNTP específico de esa reacción, lo que facilita su lectura.
En la década de 1990, la genetista Mary-Claire King demostró que un gen localizado en el cromosoma 17 era responsable de muchos casos de cáncer hereditario. Le llamó BRCA1 (por BReast CAncer gene 1). Unos años después, el médico británico Michael Stratton identificó un segundo gen implicado en el cáncer de mama hereditario localizado en el cromosoma 13 (BRCA2).
En biología, un atavismo es la reaparición de rasgos o características ancestrales que se habían perdido o suprimido durante la evolución de una especie. Por ejemplo, si un humano naciera con una cola vestigial.
Los genes dentro de una célula no funcionan de forma aislada. Se regulan unos a otros, activando o inhibiendo su expresión, formando redes complejas. El estado global de esta red (qué genes están "encendidos" o "apagados" y en qué nivel) determina el fenotipo de la célula: su tipo (neurona, célula de piel, etc.) y su comportamiento.
Es una metáfora clásica que visualiza el desarrollo y la diferenciación celular. Imagina una superficie con valles y colinas. Una célula, como una canica, rueda "cuesta abajo" por este paisaje. Las bifurcaciones en los valles representan decisiones de diferenciación, y los fondos de los valles representan los estados celulares estables y diferenciados (los distintos tipos de células maduras).